 In een cel bevinden zich nucleïnezuren. De kern bevat de grootste concentratie aan nucleïnezuren, maar ze komen ook voor in cytoplasma, mitochondriën en plastiden.
In een cel bevinden zich nucleïnezuren. De kern bevat de grootste concentratie aan nucleïnezuren, maar ze komen ook voor in cytoplasma, mitochondriën en plastiden.



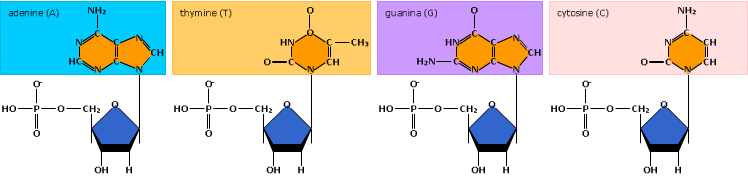
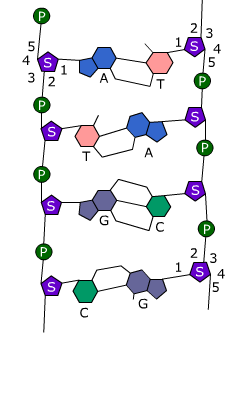 De basen van twee DNA-strengen vormen waterstofbruggen.
De basen van twee DNA-strengen vormen waterstofbruggen.
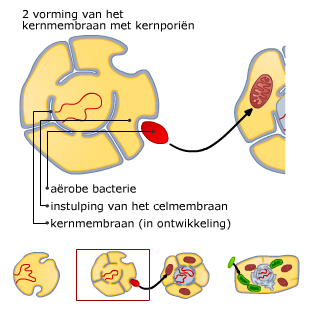
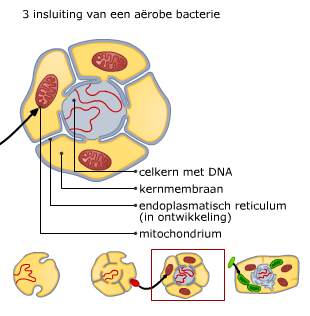
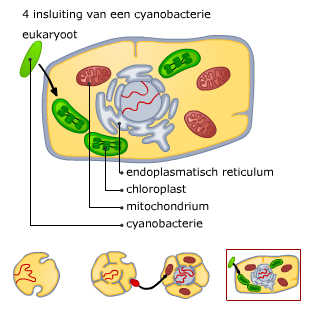
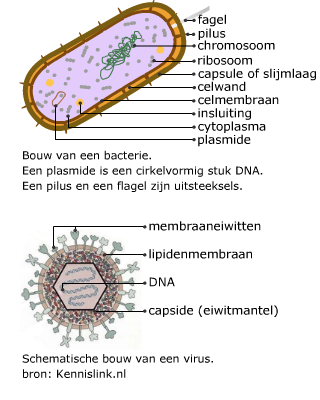 Bacteriën (prokaryoten) en virussen bevatten ook nucleïnezuren.
Bacteriën (prokaryoten) en virussen bevatten ook nucleïnezuren. Elk chromosoom van een mens bevat een DNA molecuul van ongeveer 108paren nucleotiden. Helemaal uitgestrekt zou zo’n molecuul ongeveer 4 cm lang zijn. Om alle chromosomen in een cel te laten passen en bij de celdeling over te kunnen dragen, zijn ze opgerold.
Elk chromosoom van een mens bevat een DNA molecuul van ongeveer 108paren nucleotiden. Helemaal uitgestrekt zou zo’n molecuul ongeveer 4 cm lang zijn. Om alle chromosomen in een cel te laten passen en bij de celdeling over te kunnen dragen, zijn ze opgerold.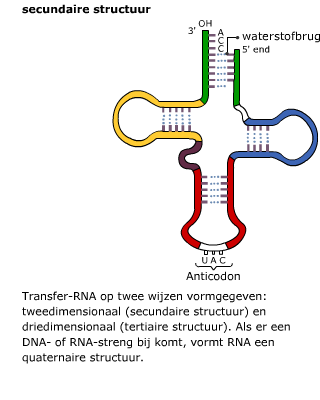
 Eiwitten spelen een onmisbare rol in het leven, als bouwstenen en als enzymen. Eiwitten kunnen zichzelf niet kopiëren; ze worden gesynthetiseerd doordat de informatie die is opgeslagen in DNA wordt vertaald.
Eiwitten spelen een onmisbare rol in het leven, als bouwstenen en als enzymen. Eiwitten kunnen zichzelf niet kopiëren; ze worden gesynthetiseerd doordat de informatie die is opgeslagen in DNA wordt vertaald.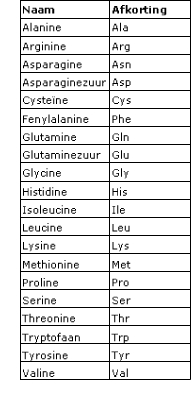 Eiwitsamenstelling
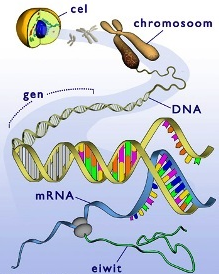
Eiwitsamenstelling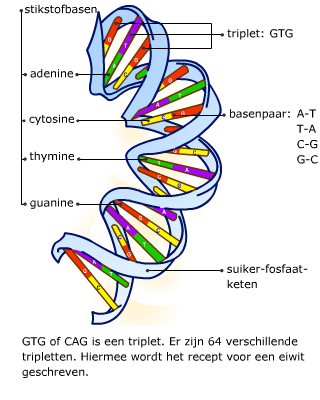 Genetische code
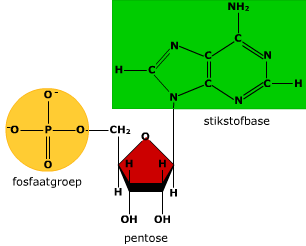
Genetische code Een stukje DNA op een chromosoom dat de informatie bevat voor één of meerdere specifieke eiwitten noem je een gen.
Een stukje DNA op een chromosoom dat de informatie bevat voor één of meerdere specifieke eiwitten noem je een gen.

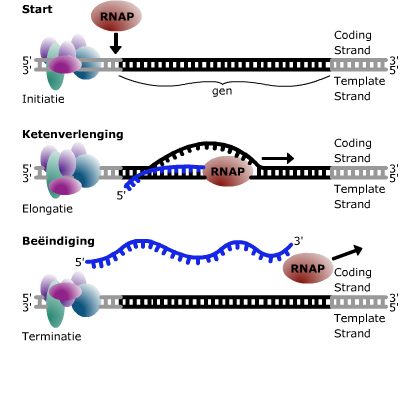
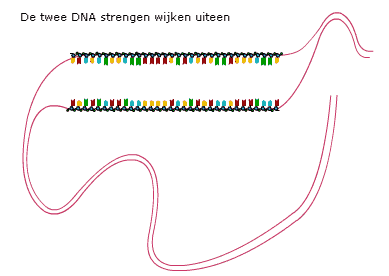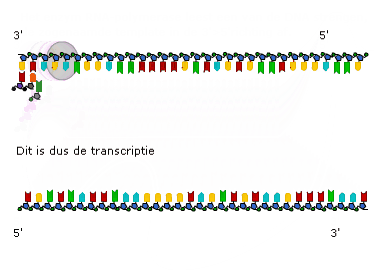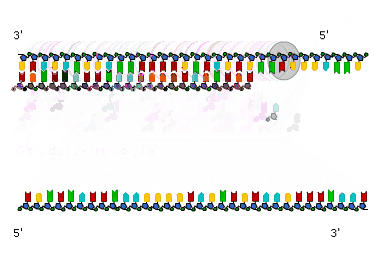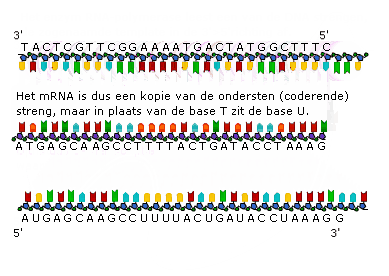 Transcriptie in detail (VWO)
Transcriptie in detail (VWO)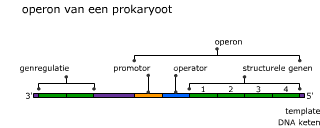 Genen die coderen voor een eiwit of voor RNA heten structuurgenen.
Genen die coderen voor een eiwit of voor RNA heten structuurgenen.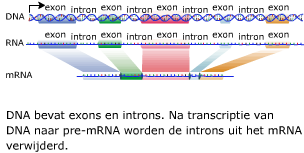 Het mRNA dat ontstaat na transcriptie (pre-mRNA) is nog niet klaar om vertaald te worden naar een eiwit. DNA bevat namelijk stukken die (waarschijnlijk) geen nuttige informatie bevatten. Deze stukken DNA heten introns. De stukken DNA die wel informatie bevatten voor het maken van een eiwit, heten exons.
Het mRNA dat ontstaat na transcriptie (pre-mRNA) is nog niet klaar om vertaald te worden naar een eiwit. DNA bevat namelijk stukken die (waarschijnlijk) geen nuttige informatie bevatten. Deze stukken DNA heten introns. De stukken DNA die wel informatie bevatten voor het maken van een eiwit, heten exons.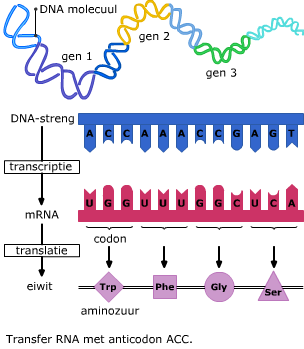 Het in elkaar zetten van eiwitten gebeurt in het cytoplasma met behulp van de ribosomen. Het mRNA beweegt zich vanuit de kern naar een ribosoom in het cytoplasma. Daar wordt de informatie op het mRNA vertaald naar een eiwit. Daarbij zijn enkele enzymen betrokken en een ander type RNA, namelijk transfer RNA ofwel tRNA.
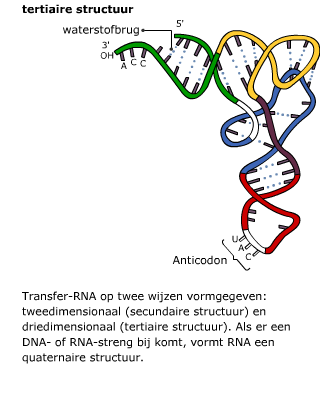
Het in elkaar zetten van eiwitten gebeurt in het cytoplasma met behulp van de ribosomen. Het mRNA beweegt zich vanuit de kern naar een ribosoom in het cytoplasma. Daar wordt de informatie op het mRNA vertaald naar een eiwit. Daarbij zijn enkele enzymen betrokken en een ander type RNA, namelijk transfer RNA ofwel tRNA.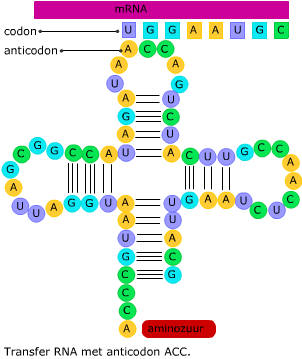 Transfer RNA bestaat uit een ruimtelijk in elkaar gevouwen “klaverblad” van RNA. Het heeft twee belangrijke bindingsplekken.
Transfer RNA bestaat uit een ruimtelijk in elkaar gevouwen “klaverblad” van RNA. Het heeft twee belangrijke bindingsplekken.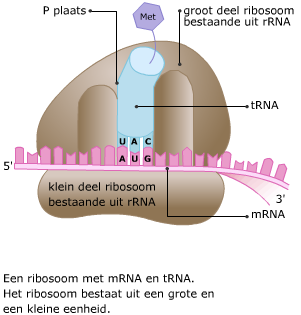 Het mRNA en het tRNA komen samen in een ribosoom, dat is opgebouwd uit eiwitten en rRNA.
Het mRNA en het tRNA komen samen in een ribosoom, dat is opgebouwd uit eiwitten en rRNA.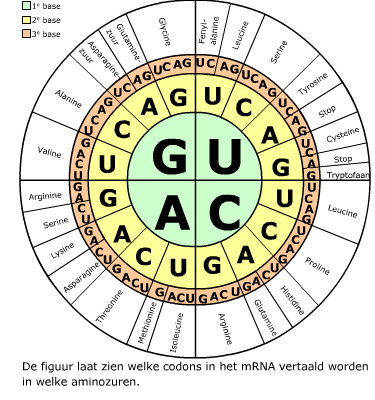 In het cytoplasma wordt de code van het mRNA vertaald in een volgorde van aminozuren. Dat proces heet translatie.
In het cytoplasma wordt de code van het mRNA vertaald in een volgorde van aminozuren. Dat proces heet translatie.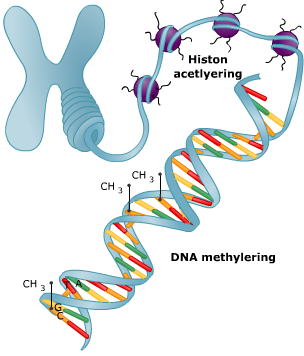 Bij prokaryoten staan genen normaal gesproken aan. Regeleiwitten kunnen bepaalde genen uitzetten.
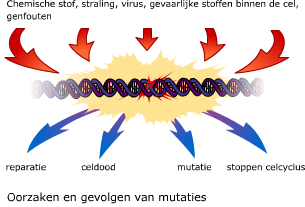
Bij prokaryoten staan genen normaal gesproken aan. Regeleiwitten kunnen bepaalde genen uitzetten.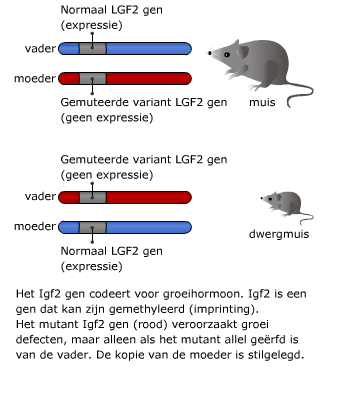 Aan cytosine van het DNA kan een methylgroep worden gebonden.
Aan cytosine van het DNA kan een methylgroep worden gebonden.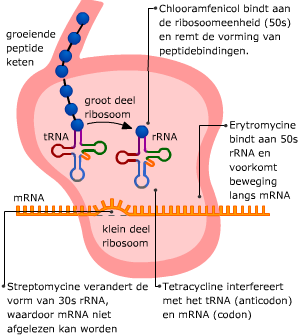 Ook na de transcriptie is dus nog regeling mogelijk.
Ook na de transcriptie is dus nog regeling mogelijk.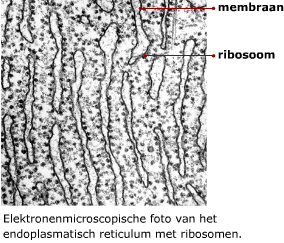 Het eiwit dat na translatie is ontstaan, wordt nog verder bewerkt, afhankelijk van de functie die het gaat vervullen. De verdere bewerking vindt plaats in het cytoplasma, in het endoplasmatisch reticulum (E.R.) of in het Golgi systeem.
Het eiwit dat na translatie is ontstaan, wordt nog verder bewerkt, afhankelijk van de functie die het gaat vervullen. De verdere bewerking vindt plaats in het cytoplasma, in het endoplasmatisch reticulum (E.R.) of in het Golgi systeem.
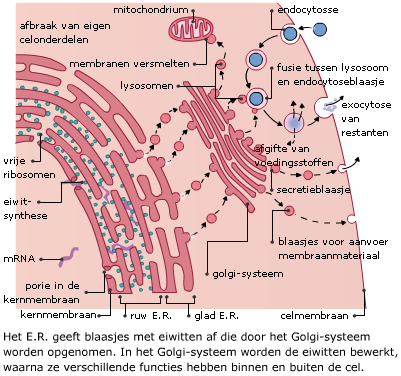 Het Golgi-systeem bevat verschillende enzymen die de in het E.R. gemaakte eiwitten verder bewerken.
Het Golgi-systeem bevat verschillende enzymen die de in het E.R. gemaakte eiwitten verder bewerken.Het arrangement Bouw en functie van nucleïnezuren is gemaakt met Wikiwijs van Kennisnet. Wikiwijs is hét onderwijsplatform waar je leermiddelen zoekt, maakt en deelt.
- Auteur
- Laatst gewijzigd
- 21-04-2021 16:08:51
- Licentie
-



Dit lesmateriaal is gepubliceerd onder de Creative Commons Naamsvermelding-GelijkDelen 4.0 Internationale licentie. Dit houdt in dat je onder de voorwaarde van naamsvermelding en publicatie onder dezelfde licentie vrij bent om:
- het werk te delen - te kopiëren, te verspreiden en door te geven via elk medium of bestandsformaat
- het werk te bewerken - te remixen, te veranderen en afgeleide werken te maken
- voor alle doeleinden, inclusief commerciële doeleinden.
Meer informatie over de CC Naamsvermelding-GelijkDelen 4.0 Internationale licentie.
Aanvullende informatie over dit lesmateriaal
Van dit lesmateriaal is de volgende aanvullende informatie beschikbaar:
- Toelichting
- De Kennisbanken bevatten de theorie bij de opdrachten.
- Leerinhoud en doelen
- Biologie;
- Eindgebruiker
- leerling/student
- Moeilijkheidsgraad
- gemiddeld